PETALURA vol. 1, 1993, p. 1
THE SKELETON-MUSCLE ORGANIZATION OF THE
HEAD FIXATION SYSTEM IN ODONATES AND ITS
EVOLUTIONARY IMPLICATIONS: A COMPARATIVE
STUDY
Stanislaw Gorb
Department of Insect Physiology
Schmalhausen Institute of Zoology
B. Chmelnickogo str. 15, Kiev
Ukraine, 252601
ABSTRACT
In imaginal odonates the head is only connected with the synthorax
through a very narrow neck region. In resting or feeding position
the dragonfly head is stabilized by a mechanical coupling of the
postcervical sclerites with specialized areas of the posterior
surface of the head capsule. In normal flight this cuppling is
inactivated and the mobile head is then acting as statolith, while
hair sensilla on its posterior side are registrating any positional
changes caused by flight manoeuvers. A single levator-muscle is
reaching from each postcervical sclerite (SPC) to a prothoracic
apodem. In flight this muscle is contracting and thus actively
abducting the SPC, that are situated within the soft cervical
membrane, to a medial position. The adduction of the SPC is achieved
passively through a spring-effect of elastic cuticular parts,
when the dragonfly is resting or feeding (in females also during
tandem-flight). In the area of contact between the SPC and the
head capsule the involved cuticular surfaces do possess significant
modifications and specializations. The adhesion in this area of
contact is supported by hypodermal secretions. This arrester-
or head fixation system is an unique character of imaginal odonates
and includes the following structures of the two body segments
head and neck:
1.) A skeleton-muscle apparatus, that moves the arrester parts.
2.) Formations with rather complicated microstructures, -
fields of microtricha on the back surface of the head (MFH) and on the postcervical
sclerites of the neck (SPC).
3.) A secretory apparatus in the hypoderma of the SPC, that
produces lipid substance into the area of contact between the
MFH and the SPC.
4.) Sensory organs (sensilles), that signalize the position
of the SPC relatively to the other neck sclerites and the posterior
surface of the head.
The MFH topography was studied in more than 100 species of
26 odonate families. Seven morphological arrester types (calopterygid,
coenagrionid, lestid, gomphid, aeshnid, anactid and libelluloid)
could be differentiated, based on peculiarities of the MFH morphology.
The skeleton-muscle organization (neck sclerites and MFH morphology,
arrester muscles and the mechanism of SPC adduction) is discussed
from a view point of constructional morphology.
The stability of the head in the SPC+MFH joint may be provided
in two ways:
1.) By enlargement of the arrester base; or ...
2.) By formation of a triangle of joints (pair of SPC + MFH
and cephaligers + tentorium).
Some evolutionary implications and conclusions are discussed.
INTRODUCTION
Fixation organs of arthropods are rather variable
in their construction and functions. An arrester- or fixation-system
of the head is unique in the animal kingdom and restricted to
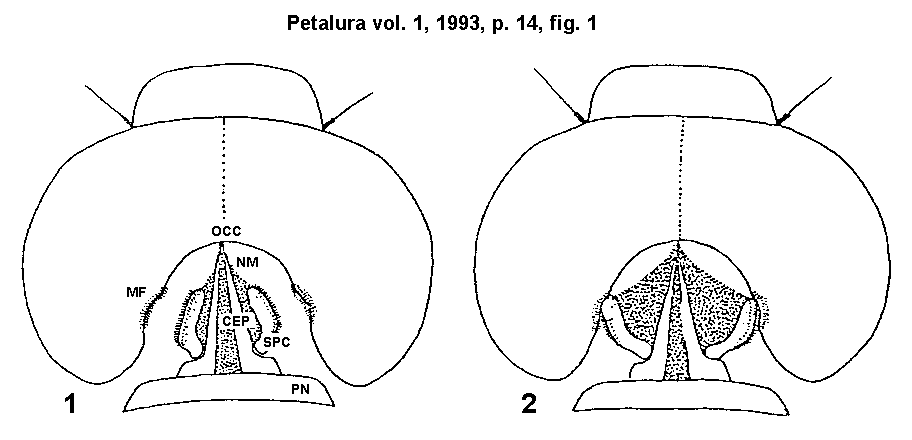
imaginal odonates (Fig. 1.).
PETALURA vol. 1, 1993, p. 2
This remarkable masterpiece of evolution includes
the following structures of the two body segments head and neck:
- 1.) A skeleton-muscle apparatus, that moves the arrester
parts.
- 2.) Formations with complicated microstructures,
such as fields of microtricha on the back surface of the head
(MFH) and on the SPC of the neck.
- 3.) A secretory apparatus in the hypoderma of the
SPC, that produces lipid substance into the region of contact
between the MFH and SPC.
- 4.) Sensory organs, that signalize the position of
the SPC relatively to the other neck sclerites and the posterior
surface of the head.
The arrester system serves for head strengthening
when the dragonfly is resting, feeding or when the female is in
tandem-position.
Morphological and physiological aspects of the neck
region in several insects, e.g. Diptera (STRAUSFELD et al., 1987), have been studied in detail, but studies in Odonata
have been scarce. Four cervical sclerites of imaginal Libellula
auripennis Burm. (Libellulidae) and two of imaginal Lestes
uncata Kirby (= L. dryas Kirby) (Lestidae) were illustrated
by SNODGRASS (1909). A comparison of neck
sclerites in different insects, including a larval Uropetala
(Petaluridae) and an imaginal Epiophlebia superstes Selys
(Epiophlebiidae) was briefly described and figured by CRAMPTON (1926). The nomenclature of neck sclerites after SNODGRASS and CRAMPTON with original names was given
by ASAHINA (1954). His table included data
for six odonate taxa: Anisozygoptera: Epiophlebia (Epiophlebiidae);
Zygoptera: Mnais (Calopterygidae); Anisoptera: Orthetrum
(Libellulidae), Gynacantha (Aeshnidae), Davidius
(Gomphidae) and Tanypteryx (Petaluridae). The functional
morphology of the neck in feeding larval and imaginal Aeshna
juncea L. (Aeshnidae) was investigated by POPHAM
& BEVANS (1979). The general morphology
of the dragonfly head capsule was studied by STEINMANN
(1967) and ASAHINA (1957). The function of
the MFH and SPC as joint organ was proposed for the first time
by BERLESE (1909). He suggested that these
structures might be organs for stridulation ("laminae stridulatoriae").
TILLYARD (1917) doubted this function and
a head-holder function of these organs ("Kopfklemmen")
was briefly discussed by MITTELSTAEDT (1950)
in his work about dragonfly flight reflexes. This hypothesis of
Mittelstaedt has been supported by PRINGLE
(1963).
The dragonfly arrester system has been especially
investigated in the fields of external and internal morphology
(GORB, 1988; 1989; 1990a: 1990b; 1991a), histology
(GORB, 1990b) and physiology (GORB,
1991b; 1991c). Nevertheless some questions have not been treated
in these papers: Distinctions of muscle topography in several
taxa; the mechanism of SPC adduction; correlation between size
and topography of the MFH with the morphology of the head capsule,
in different taxa. In the present paper I am discussing these
questions, and also peculiarities of the skeleton-muscle organization
of the arrester system, from a viewpoint of constructional morphology,
together with some evolutionary considerations.
PETALURA vol. 1, 1993, p. 3
SPECIES EXAMINED
The external morphology of more than 100 species
of 26 families of the order Odonata has been examined:
Hemiphlebia mirabilis Selys (Hemiphlebiidae);
Synlestes weyersi Selys (Chlorolestidae);
Lestes barbarus Fabricius, L. dryas Kirby,
L. sponsa Hans., L. virens Charp. (Lestidae);
Argiolestes minimus Tillyard, Dimeragrion
percubitale Calvert, Heteragrion chrysops
Hagen in Selys, Oxystigma williamsoni Geijskes,
Philogenia cassandra Hagen in Selys (Megapodagrionidae);
Palaemnema clementina Selys (Platystictidae);
Mecistogaster linearis Fabricius, Megaloprepus
caerulatus Drury, Pseudostigma aberrans Selys
(Pseudostigmatidae); Platycnemis pennipes
Pall. (Platycnemididae); Argia vivida Hagen
in Selys, Coenagrion pulchelum Van der Lind., Erythromma
najas Hans., Enallagma cyathigerum Charp.,
Ischnura elegans Van der Lind. (Coenagrionidae);
Isosticta spinipes Selys (Isostictidae);
Nososticta solida Hagen in Selys, Prodaisineura
vittata Selys, Epipleoneura lamina Williamson,
Psaironeura remissa Calvert (Protoneuridae);
Diphlebia lestoides Selys (Diphlebiidae);
Rimanella arcana Needham (Amphipterygidae);
Libellago lineata Burmeister, Rhinocypha fenestrella
Rambur (Chlorocyphidae); Bayadera indica
Selys, Euphaea variegata Rambur (Euphaeidae);
Euthore fasciata Hagen in Selys, Cora cyane
Selys, Chalcopteryx rutilans Rambur, Polythore
ornata Selys (Polythoridae); Heliocharis
amazona Selys (Dicteriadidae); Iridictyon
trebbaui Racenis (Calopterygidae); Epiophlebia
superstes Selys (Epiophlebiidae); Petalura gigantea
Leach, Phenes raptor Rambur, Tachopteryx
thoreyi Hagen, Uropetala carovei White, Tanypteryx
pryeri Selys (Petaluridae); Aeshna affinis
Van der Lind., Ae. mixta Latr., Ae. grandis L.,
Anaciaeschna isosceles Müll., Anax goliath
Selys, A. imperator Leach, Gynacantha nervosa
Rambur, Staurophlebia reticulata Burm., Polycanthagyna
melanictera Selys, Acanthaeschna victoria
Martin, Tricanthagyna trifida Rambur, Aeschnophlebia
anisoptera Selys, Austroaeschna multipunctata Martin,
Brachytron pratense Müll., Boyeria vinosa
Say, Oligoaeschna modiglianii Selys (Aeshnidae);
Stylogomphus suzukii Matsumura in Oguma, Sinogomphus
flavolimbatus Oguma, Aphylla producta Selys,
Cyclophylla breviphylla Belle, Phyllogomphoides
sp., Progomphus obscurus Rambur, Zonophora
batesi Selys, Sieboldius albardae Selys,
Gomphidia kruegeri Martin, Ictinogomphus
acutus Laidlaw (Gomphidae); Chlorogomphus
magnificius Selys, Cordulegaster annulatus
Don., Anotogaster sieboldii kuchenbeiseri
Foerster, Zoraena diastatops Selys (Cordulegastridae);
Eusynthemis brevistyla Selys (Synthemistidae);
Epophthalmia vittigera Rambur (Macromiidae);
Cordulephya pygmaea Selys, Oxygastra cutisii
Dale, Gomphomacromia paradoxa Brauer, Somatochlora
flavomaculata Van der Lind., Epitheca bimaculata
Charp., Cordulia aenea L. (Corduliidae);
Anatya anomala Kirby, Brachidiplax sobrina Rambur,
Uracis imbuta Burm., Celithemis eponina Drury, Leucorrhinia
pectoralis Charp., Libellula depressa L., L. quadrimaculata
L., Orthetrum cancellatum L., Sympetrum flaveolum
L., S. meridionale Selys, S. vulgatum L.,
Dythemis velox Hagen, Onychothemis abnormis Brauer,
Diastatops pullata Burm., Zenitoptera americana L.,
Camacinia gigantea Brauer, Tramea chinensis Burm.,
Zyxomma petiolatum Rambur, Celebothemis delecollei Ris,
Zygonyx iris Selys, Aethriamanta brevipennis Rambur
(Libellulidae).
As comparative material, specimens of other insect
orders, such as Neuroptera (Nemoptera sp., Myrmecoleon
sp., Ascalaphus libelluloides), Mecoptera (Panorpa
communis) and Embioptera (Haploembia
solieri), have been studied too.
METHODS
1.) Light microscopy: Whole-mount preparations of
the MFH and SPC, embedded in canada-balsam, have been examined,
drawed and photographed with a binocular microscope MBI-11 (magnifications:
*200, *400, *600).
2.) Scanning electron microscopy (SEM): The MFH and
neck region were air-dried, mounted on holders, sputter-coated
with gold and examined with the scanning electron microscopes
JEOL JSM-35C, JEOL JSM-T20 and TESLA-BS-301.
3.) Morphometry: 12 values of the head in 26 species
of 6 families have been measured (Fig. 2.). In all cases 3-7 specimens
of each species have been examined. According to STEINMANN
(1967) the values of the dragonfly head are sufficiently constant
to warrant such a morphometric analysis.
PETALURA vol. 1, 1993, p. 4
The measurement pattern adequately described
the general characters of the head capsule as well as the size
and topography of the cephalic fields of microtrichia. Correlation
coefficients for every pair of values of Lestes- and Sympetrum-samples
have been calculated with a personal computer, and clusters according
to the closest-neighbour-method were created.
4.) Abbreviations: BS - basisternum, CEP - cephaliger,
CX - coxa, LM (or M1) - abductor muscle of the postcervical sclerite,
MFH - microtrichia field of the head, NM - neck membrane, OCC
- occiput, PN - pronotum, SEC - eucervical sclerit, SPC - postcervical
sclerit, TR - tentorium.
RESULTS
1.) Neck sclerites morphology: The dragonfly imago
has a well developed neck region. The lateral walls of the neck
are built by a pair of eucervical sclerites (Fig. 3.). These
SEC are symmetrical relatively to the longitudinal body axis and
are anteriorly transformed into two acute processes, called cephaligers.
The cephaligers converge towards the centre of gravity of the
head. In this point the head tentorium connects with the neck
through a pair of mobile joints (Fig. 4.). Ventrally the SEC connect
with the basisternum through the neck membrane. The microsculpture
of this neck membrane is rather complicated and differs in representatives
of different families. In the CEP-region the SEC connects with
the neck membrane, which provides mobility for the SEC. The SEC
can be moved by a neck muscle system over an axis that crosses
the dorso- and ventro-proximal edges of the sclerites. Near their
dorso- proximal edge the SEC are connected with the postcervical
sclerites. They are isolated from the SEC and connect with NM
from three sides. Such an articulation promotes high mobility
of the SPC. The SPC are differentiated into a sclerotized basis
and an elastic distal part. The distal part is rounded or elongated,
and covered by microtrichia, by hair and campaniform sensilles.
2.) Arrester muscles: The SPC is abducted by a SPC-levator-muscle
(LM). Its origin is an apodem of the pronotum, its insertion
is the cuticular region between the SEC and the SPC. The topography
of the muscle insertion was similar in all studied specimens.
The pronotal apodem is differently developed and successively
elongated in the sequence Cordulegaster, Sympetrum,
Cordulia, Aeshna and Gomphus. Consequently
the topography of the muscle origin changes too. The
directions of forces relatively to the SPC, caused by the LM,
are different in different taxa. The LM may be directed postero-dorso-medially
(Sympetrum, Cordulegaster, Lestes), dorso-medially
(Aeshna, Cordulia, Calopteryx) or anterio-dorso-medially
(Gomphus) in species with a long pronotal apodem (Fig. 3.).
As soon as the abduction of the SPC to the longitudinal
body axis (by the LM) is completed, the sclerites (SPC) do not
contact anymore with the MFH, consequently the head is free and
mobile. To establish the SPC-MFH contact, the head must be closer
to the neck. The head-neck muscle system, which is similar in
most insects, is achieving this effect.
PETALURA vol.1, 1993, p. 5
This muscular complex consists of three pairs
of muscles that are moving the head in the horizontal plane (yaw),
sagittal plane (roll) and transversal plane (pitch) (MITTELSTAEDT,
1950; PRINGLE, 1963; STRAUSFELD
et al., 1987).
3.) The mechanism of SPC adduction: An antagonistic
muscle to the SPC - abductor has not been found. The sclerit adduction
is passively established through the elasticity of peculiarly
organized cuticular structures. The SPC and SEC are posteriorly
connected by an elastic cuticular part and are moved independently.
This cuticular part can adduct the SPC like a spring. The spring-effect
is increased by a pair of hard cuticular "legs" of the
SPC-SEC region in Calopteryx, Lestes, Sympetrum,
Gomphus and Cordulegaster (Fig. 5.). When the SPC
are abducted by the LM, these "legs" are twisting. When
the LM are relaxing, the "legs" are untwisting and adduct
the SPC. Besides, the SPC-"legs" of Sympetrum
do have two supplementary parts of elastic cuticula, - posteriorly
and anteriorly. They are probably also involved in sclerit adduction.
A soft and elastic cuticula of the distal part of the SPC is damping
external forces and stresses on the head capsule, when the SPC
is connecting with the MFH.
4.) MFH topography: The MFH and SPC are functionaly
connected structures and are consequently correlated in form,
size and type of microtrichia. On the other hand some great differences
in MFH size and topography have been noticed between the different
odonate families, which are probably correlated with several pecularities
of the head capsule.
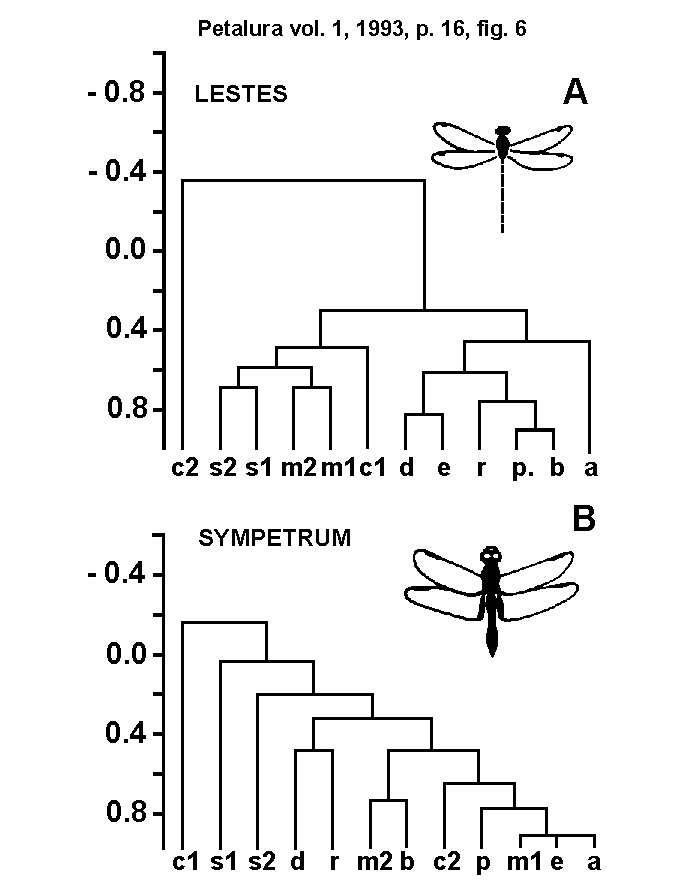
- 4.1.) Cluster analysis: To test this hypothesis,
morphometric characters of the head capsule and the MFH of Sympetrum
(Libellulidae) and Lestes (Lestidae) have been studied,
using the closest-neighbour method of cluster analysis. These
two taxa were selected because of their clearly isolated and more
or less rounded MFH, that can be easily measured, and because
their head-capsule and MFH morphology is sufficiently variable.
For the cluster analysis 4 or 5 specimens of each of the following
species have been examined: Lestes barbara, L. dryas,
L. sponsa, L. virens and Sympetrum
meridionale, S. flaveolum, S. sanguineum, S.
vulgatum. The resulting clusters are shown in Fig. 6. The
character-distribution in the clusters of Lestes differs
from those of Sympetrum. For the characters of Lestes
two clearly differentiated groups could be observed. The first
group includes characters of length, width and height of the head
capsule (A, B, R, D, E, P); while the second group includes characters
of the MFH size (S1, S2) and of the occiput position in dorso-ventral
direction. Thus the MFH size does better correlate with the latter
characters than with those of the head capsule size and the characters
correlated with the position of the MFH in medio-lateral direction.
The character-distribution of the Sympetrum clusters is
differing in its almost hierarchical distribution of character
correlation. Here the MFH size is less correlated with other characters
of the head capsule, but with head width and length. The MFH size
(S1, S2) is in first place correlated with head length (D, R).
Another mathematical analysis showed that the position
of the fixation joints relatively to the foramen magnum (= sagittal
turning axis of the head) has the least variation among twelve
characters of the head fixation system (GORB,
1991c).
PETALURA vol.1, 1993, p. 6
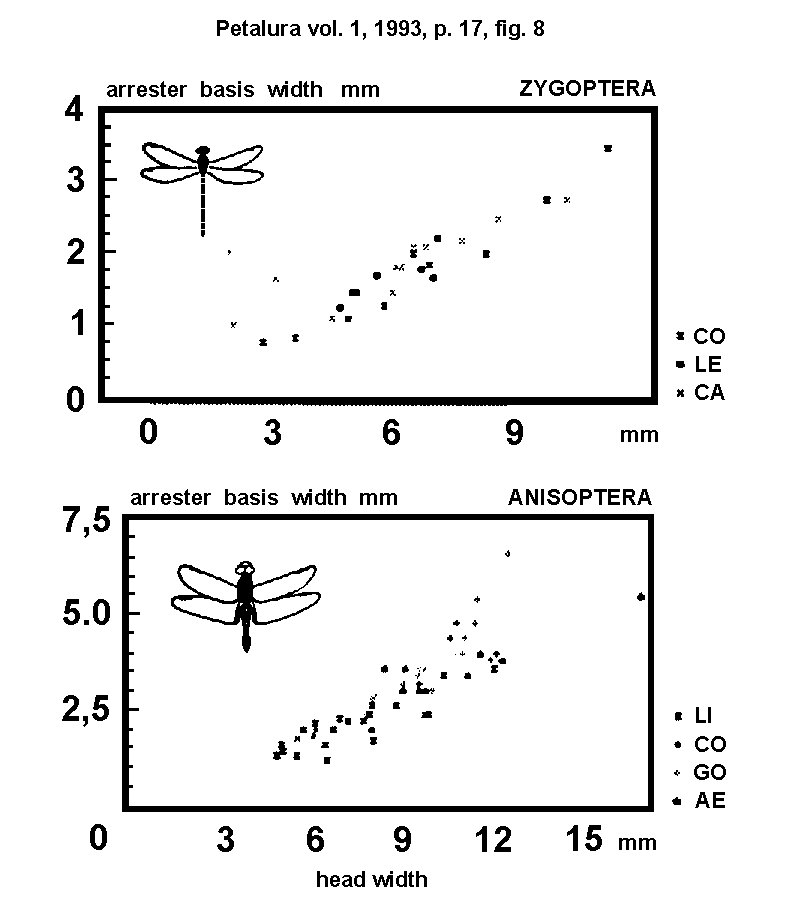
- 4.2.) Arrester base: The relation of maximal head
length to the base length of the head-neck junction (= maximal
width of the occiput) in insects is usualy varying between 2 :
1 and 4 : 1. In odonates this relation is varying between 8 :
1 (Petaluridae) and 16 : 1 (Hemiphlebiidae). A small base of
the head-neck junction causes a weak mechanical stability. When
the head is fixed to the neck at the MFH points, the base of the
head-fastening is enlarged, because in this state the base of
the arrester-system (= the distance between right and left points
of fixation) is serving as base of head-fastening. This mechanism
is providing a better stability of the head. The cluster distribution
is showing that the arrester base (B) does better correlate with
the MFH position in dorso-ventral direction (Lestes) and
with the size of the mouthparts (Sympetrum), than with
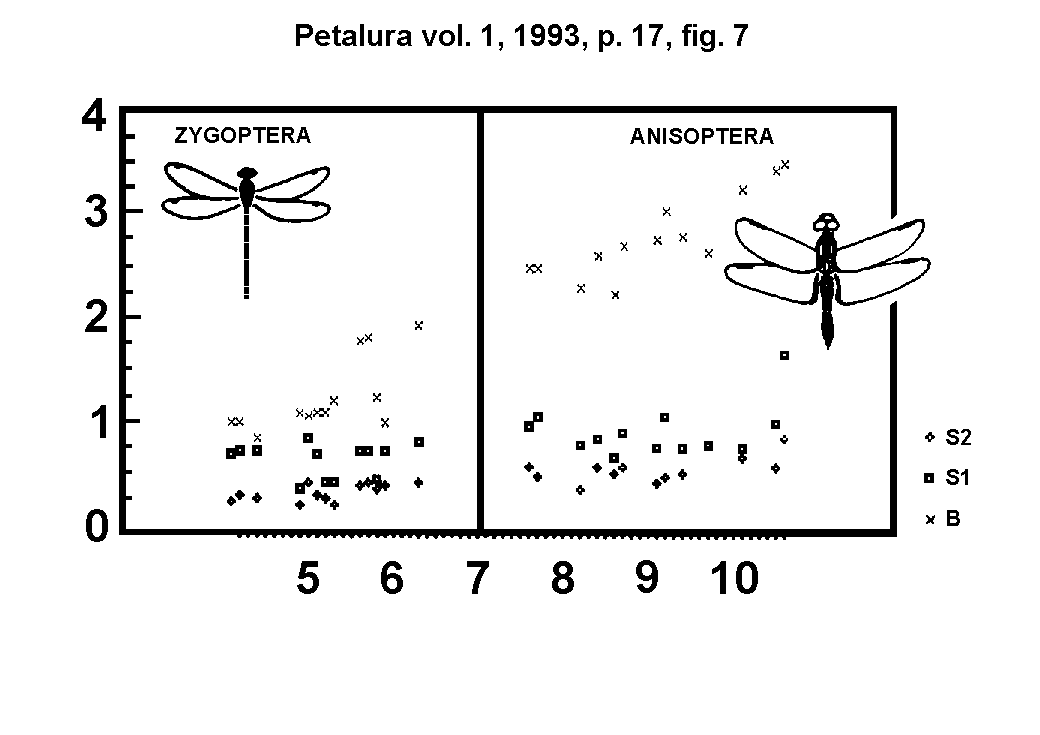
the head width. The dependance of the arrester base from the head
width is linear for the six odonate superfamilies (Fig. 7.). Besides,
the angles between these lines and the coordinate-lines are different
for the different odonate families and superfamilies. This fact
can be explained by the constructional differences of the head
capsule (compare Aeshnidae with Gomphidae and Libelluloidea, or
Coenagrionoidea with Calopterygoidea).
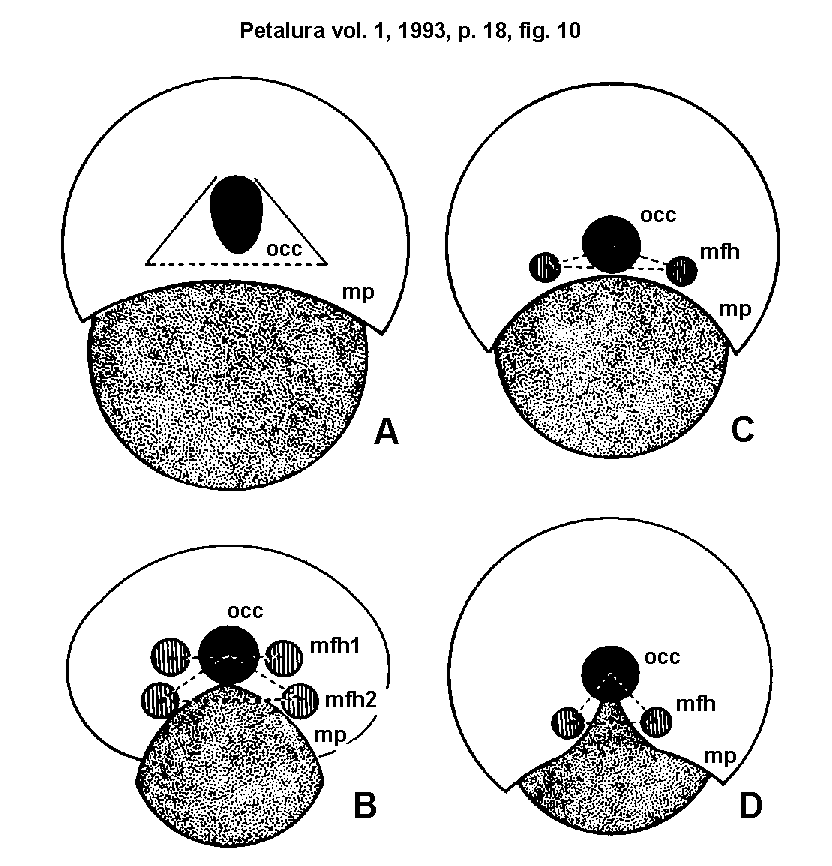
5.) Morphological types of the MFH:
The different types of fixation systems are nearly always family-characteristic, rarely genus- or species-characteristic.
PETALURA vol.1, 1993, p. 7
ANISOPTERA:
- d) Gomphid type: The shape of the head capsule is
varying from medio-laterally widened (Epiophlebia, Gomphus,
Sinogomphus) to dorso-ventrally heightened (Phenes,
Uropetala). Laterally of the occiput there is a cuticular
surface, sparsely covered with microtrichia and hair sensilles.
Isolated MFH are absent. The cuticular fold extends ventro-laterally
from the dorsal edge of the occiput. The SPC contacts with this
fold, which seems to be a peculiar slot for the SPC. The gomphid
type is characteristical for the Epiophlebiidae, Petaluridae and
Gomphidae.
- e) Aeshnid type: The head capsule is nearly ellipsoidal
and medio-laterally widened. There is one clearly isolated MFH
on each side, laterally of the occiput on the level of its ventral
edge. The microtrichia of the MFH are very large and spine-like; in aeshnids they do possess small canals. This type is
characteristical for the Aeshnidae (except Anactini) and also
for a part of the Cordulegastridae (the Cordulegastrinae seem
to be intermediate between the aeshnid- and the libelluloid-type,
while the Chlorogomphinae seem to be intermediate between the
gomphid-type and the type present in cordulegastrines).
- f) Anactid type: The head capsule is of the same
shape as that of other aeshnids, but there are two MFH on each
side of the head. This type is only present in representatives
of the aeshnid tribe Anactini.
- g) Libelluloid type: The head capsule is ellipsoidal
(ovoidal) (Tramea, Zyxomma, Cordulephya)
or slightly medio-laterally widened (Celebothemis, Palpopleura,
Onychothemis). There is only one clearly isolated MFH on
each side of the head, situated lower than the ventral edge of
the occiput. This type is characteristical for all Libelluloidea
(Synthemistidae, Macromiidae, Corduliidae and Libellulidae).
DISCUSSION
Imaginal dragonflies do possess a well developed
neck region compared to dragonfly larvae or other insect orders
(CRAMPTON, 1926; POPHAM
& BEVANS, 1979). A good mobility
of the head is necessary for the signalization of body-turns when
the dragonfly is in high-speed and intensively manoevering flight
(MITTELSTAEDT, 1950). The start of flight
muscle activity is inducing the inactivation of the head fixation
by contraction of the SPC-levator-muscle. The head is acting as
statolith and contacts with hairy plates of the neck and pronotum.
This mobility of the head is a result of reducing the base of
the head-neck joint nearly to a single point. Consequently this
articulation does only have a weak mechanical stability. In several
behavioural situations the arrester system can eliminate this
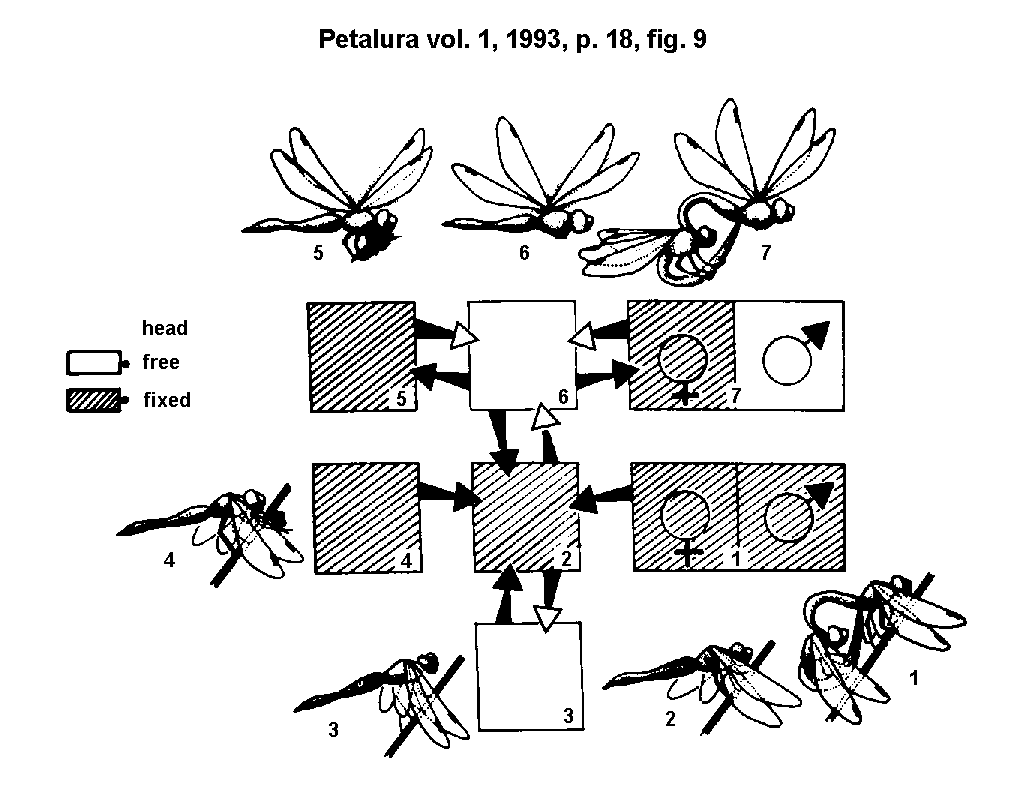
shortcoming by enlarging this base (GORB,
1991b; 1991c) (Fig. 9.). Sensory controll of the head fixation
is achieved by trichoid sensilla of the head capsule and campaniform
sensilla of the SPC. The phenomenon of head fixation
is an exceptional case among arthropods. The only other case described
in the entomological literature is the head fixation of the Isoptera,
managed by mandibular processes that fit into hollows of the prothorax.
PETALURA vol. 1, 1993, p. 8
This kind of fixation is increasing the stability
of the head-prothorax articulation when the mouthparts are cutting
a plant stem (ILYCHEV, 1987).
The odonate arrester system is unique because of
the following features:
- 1.) Absence of antagonistic muscles: Muscular movements
without antagonistic muscles have only been found in some arthropods.
Usually such an absence of antagonistic muscles is compensated
by hydraulic mechanisms, e.g. in the odonate larval mask (OLESEN,
1972; 1979; TANAKA & HISADA,
1980) and spider legs (FOELIX, 1982), or by
cuticular springs, e.g. in the cranio-mandibular system of ants
of the tribes Dacetini and Odontomachini (DLUSSKY
& FEDOSEEVA, 1988).
- 2.) Different modifications of the type of fixation:
- a) The well developed surface of the SPC and MFH,
covered with fine microtrichia and accompanied by hypodermal secretion.
This type is present in the majority of odonate taxa.
- b) SPC with hook-like microtrichia and MFH with
mushroom-like microtrichia. This type is present in the lestid
genus Sympecma.
- c) SPC with club-like microtrichia, while the MFH
acts as elastic hinge. This type is present in the lestid genus
Lestes.
The structure of the microtrichia on head and neck
is an important morphological base for the type of fixation. The
presence of different types of microtrichia and consequently of
different types of fixation, is explaining the poor correlation
of head shape and size with the MFH size. The stability of adhesion
is achieved by different principles of fixation (different microtrichia
types), and not by enlargement of the area of contact. Important
factors for this stability also seem to be the secretion at the
SPC-region and the correlation between the microtrichial density
on the SPC and MFH (GORB, 1990b).
Comparing rather closely related genera like Aeshna
and Anax or Lestes and Coenagrion, one will
notice that only a single MFH is present in Aeshna and
Lestes, while two MFH of identical size are present in
Anax and Coenagrion, on each side of the posterior
surface of the head respectively. Double MFH do provide an enlargement
of the area of adhesion with the SPC. This type of arrester system
allows two possible positions for the SPC (vental and dorsal).
Probably intermediate positions for the SPC are also possible,
between the ventral and dorsal MFH. The long and flatened microtrichia
of the dorso-ventally heightened SPC of gomphids do contact directly
with the smooth posterior surface of the head, because gomphids
do not possess any MFH. This type of fixation is working just
like the fixation of a surface with fine microtrichia plus hypodermal
secretion in the praetarsus of many insects, such as Hemiptera,
Coleoptera, Mecoptera and Diptera (HASENFUSS,
1978; BAUCHENß, 1979; BAUCHENß
& RENNER, 1977; WIGGELSWORTH,
1987; RÖDER, 1984; 1986; GHASI-BAYAT,
1979; GHASI-BAYAT &
HASENFUSS, 1980a; 1980b; STORK,
1980; 1981a; 1981b; 1983). By analogy with some Orthoptera (BERNAYS,
1986; BERNAYS & HAMAL,
1987) we may suppose that different forms of head capsules are
diet-induced. Different types of head capsule and probably also
different types of chasing behaviour (ZAIKA
& VORONOVA, 1977; GORB,
1991b) resulted in different constructional types of the arrester
system.
PETALURA vol. 1, 1993, p. 9
From a viewpoint of constructional morphology there
are several constraints concerning the MFH-topography (BAREL
& ANKER et al., 1989). To improove head
stability during head fixation, a maximum increase of the arrester
base is necessary, which could be achieved by a lateral shift
of the MFH; but in this case the SPC and its muscles would be
essentially lengthened and SPC adduction could not be achieved
by cuticular springs. Another constructional constraint, especially
in Zygoptera, seems to be the eye-size. The most important constraint
is the topographic position of the MFH on the dorso-ventral axis,
in relation to the occiput. If the MFH is situated lower than
the occiput, then the system in fixed position does have a maximum
stability, because the fixation points are forming a triangle
(left MFH - CEP - right MFH). This kind of MFH position is only
present in some evolutionary advanced taxa, such as Coenagrionoidea,
Anax and some libellulids. Most other taxa do have an extremely
ventrally placed MFH, which is often even situated close to the
hypostomal suture. In contrast to the triangle formation, there
is some tangential mobility when the head arrester system is in
"fixed" position. In the gomphid-type of arrester system
there are other reasons for a free mobility of the head in "fixed"
position. In this particular case the arrester construction allows
shifting of the SPC along the cuticular fold on the posterior
surface of the head capsule. In all these types of arrester system
the most important constructional constraints for the dorso-ventral
position of the MFH are the size and topography of the mouthparts.
The mouthparts of aeshnids and especially of petalurids are somewhat
larger than those of libellulids. In taxa with relatively larger
mouthparts the extreme ventral position of the MFH is on the level
of the ventral edge of the occiput, close to the hypostomal suture.
When the MFH are situated considerably lower than the occiput,
the arrester base is smaller than it is when the MFH are situated
on the same level as the occiput (Fig. 9.; compare the arrester
base of libellulids with gomphids and aeshnids for the same values
of head width).
Consequently the stability of the arrester joint
can be achieved in two different ways:
- 1.) By increase of the arrester base.
- 2.) By formation of a triangle of joints.
This second type is evolutionary younger and characteristical
for the Libelluloidea.
Evolution of the fixation mechanisms and cuticular
structures:
- a) Elastic "hinge"-type: Present in the
Chlorolestidae and Lestinae of the Lestinoidea.
- b) hook-type: Present in the evolutionary advanced
taxa Sympecmatinae (Lestidae), Coenagrionoidea and Libelluloidea.
- c) "Pulvilles"-type with greatly increased
surface: Present in the Aeshnidae and Gomphidae etc.
The general character distribution of the fixation
system is rather congruent with the "traditional" phylogenetic
tree of odonates. The Epiophlebiidae ("Anisozygoptera")
did retain the highest number of plesiomorphic characters. Within
the Anisoptera petalurids and gomphids have preserved many plesiomorphic
characters. Within zygopteres there are two different evolutionary
directions:
- a) Diminution of the microtrichial fields on the
head capsule (MFH) in Lestinoidea.
- b) Development of two microtrichial fields on each
side of the head.
PETALURA vol. 1, 1993, p. 10
ACKNOWLEDGEMENTS
Prof. Dr. L.I. FRANTSEVICH, Dr. P.A. MOKRUSHOV, Dr.
V.N. FURSOV, S.S. KULCHITSKY (Schmalhausen Institute of Zoology,
Kiev, Ukraine); Dr. L.N. PRITYKINA (Palaeontological Institute,
Moscow, Russia); Dr. L.A. ZHILTSOVA (Zoological Institute, Sankt-Petersburg,
Russia); Prof. Dr. B.F. BELYSHEV, Dr. Yu. HARITONOV (Biological
Institute, Novosibirsk, Russia); Dr. S.N. BORISOV (Institute of
Zoology and Parasitology, Dushanbe, Tadjikistan); Dr. R.S. PAVLJUK
(Museum of Zoology, Lwiw University, Ukraine); Prof. Dr. N.N.
SHCHERBAK (Museum of Zoology, Kiev, Ukraine); Dr. J. DE MARMELS
(Universidade Central de Venezuela); Prof. Dr. M. SAMWAYS (Univerity
of Natal, South Africa); Mr. M. SUGIMURA (Dragonfly Museum, Nakamura,
Japan); Dr. M.M. EIDELBERG (Nikitsky Botanical Garden, Jalta,
Ukraine); kindly gave me specimens of different taxa. I also thank
V.A. KRIVOSHEEV (Botanical Institute, Kiev) and N.T. STRASHKO
for their assistance in SEM-microscopy, B.O. MIKHALEVICH for original
BASIC-programms for cluster analysis, E.V. GORB (University of
Kiev) for helping with the translation of this manuscript, and
an anonymous referee. I am very grateful to Mr. G. BECHLY (University
of Tübingen, Germany) for his advices and assistance with
this paper.
REFERENCES
ASAHINA, S. (1954): A morphological study of a relic dragonfly Epiophlebia
superstes Selys (Odonata: Anisozygoptera). The Japan Society
for the Promotion of Science: Tokyo. iV + 154 pp., 71 pl.
BAREL, C.D.N. & ANKER,
G.Ch. et al. (1989): Constructional constraint
and its ecomorphological implications. Acta Morphol. Neerl.-Scand.,
27: 83-109
BAUCHHENß, E. (1979): Die Pulvillen von Calliphora erythrocephala Meig.
(Diptera: Brachycera) als Adhäsionsorgane. Zoomorphologie,
93(2): 99-123
BAUCHHENß, E. &
RENNER, M. (1977):
Pulvillus of Calliphora erythrocephala Meig. (Diptera: Calliphoridae).
Int. J. Insect Morphol. and Embryol., 6(3-4): 225-227
BERLESE, A. (1909): Gli Insetti. Vol. 1. Milano. 1004 pp.
BERNAYS, E.E. (1986): Diet induced head allometry among foliage-chewing insects
and its importance for graminivores. Science, 231(4737):
495-497
BERNAYS, E.E. & HAMAI, (1987): Head size and shape in relation to grass feeding in Acridoidea
(Orthoptera). Int. J. Insect Morphol. and Embryol., 16(5-6):
323-330
CRAMPTON, G.C. (1926): A comparison of the neck and prothoracic sclerites throughout
the orders of insects. Trans. Amer. Ent. Soc., 52:
192-248
ILYCHEV, V.D. (ed.) (1987): BIOLOGICAL DAMAGES. The Manual for biological faculties
of a High School. High School: Moscow. 352 pp. (in russian)
PETALURA vol. 1, 1993, p. 11
DLUSSKY, G.M. & FEDOSEEVA,
E.B. (1988): Origin and early evolution
of ants (Hymenoptera: Formicidae). p. 16-62 in: Cretaceous
biocenotic crisis and insect evolution. Nauka: Moscow. (in russian)
FOELIX, R.F. (1982): The Biology of Spiders. Harvard University Press:
Massachusetts and London. 306 pp.
GHASI-BAYAT,
A. (1979): Zur Oberflächenstruktur
der tarsalen Haftlappen von Coreus marginatus (L.) (Coreidae,
Heteroptera). Zool. Anz., 203(5/6): 345-347
GHASI-BAYAT,
A. & HASENFUSS, I. (1980a): Über den Transportweg der Haftflüssigkeit der
Pulvilli bei Coptosoma scutellatum (Geoffr.) (Plataspidae,
Heteroptera). Nachrichtenbl. Bayer. Entomol., N 1:
58
GHASI-BAYAT,
A. & HASENFUSS, I. (1980b): Zur Herkunft der Adhäsionsflüssigkeit der tarsalen
Haftlappen bei den Pentatomidae (Heteroptera). Zool. Anz.,
204(1/2): 13-18
GHASI-BAYAT,
A. & HASENFUSS, I. (1980c): Die Oberflächenstrukturen des Prätarsus von
Elasmucha ferrugata (Fabricius) (Acanthosomatidae, Heteroptera).
Zool. Anz., 205(1/2): 76-80
GORB, S.N. (1989): Functional morphology of the arrester system in dragonflies.
Vestn. zool., N 3: 62-67. (in russian)
GORB, S.N. (1990a): The microsculpture of the head fixation system in dragonflies
in a scanning electron microscope. Zool. zh., 69(2):
148-154. (in russian)
GORB, S.N. (1990b): The inner morphology of the arrester system in the dragonfly
Erythromma najas Hansemann (Zygoptera: Coenagrionidae).
Vestn. zool., N 6: 59-62+1. (in russian)
GORB, S.N. (1990c): The external morphology of the head fixation system in
the dragonflies of the superfamily Calopterygoidea (Odonata: Zygoptera).
Zool. zh., 69(11): 37-45. (in russian)
GORB, S.N. (1991a): Sensilla in the arrester system of dragonflies (Odonata).
Vestn. zool., N 1: 61-68. (in russian)
GORB, S.N. (1991b): Head fixation reflexes in flight of dragonflies. Zh.
Evol. Biokh. Physiol., 27(4): 46-51. (in russian)
GORB, S.N. (1991c): Functional morphology of the arrester system in dragonflies
(Odonata). Ph.D.-thesis, Zoological Institute Kiev. 403 pp.
(in russian)
HASENFUSS, I. (1978): Über das Haften von Insekten an glatten Flächen
- Herkunft der Adhäsionsflüssigkeit. Zool. Jahrb.,
Abt. 2, 99(1): 115-116
MITTELSTAEDT, H. (1950): Physiologie des Gleichgewichtssinnes bei fliegenden Libellen.
Z. vergl. Physiol., 32(5): 442-463
OLESEN, J. (1972): The hydraulic mechanism of labial extension and jet propulsion
in dragonfly nymphs. J. Comp. Physiol., 81(1): 53-56
OLESEN, J. (1979): Prey capture in dragonfly nymphs (Odonata; Insecta): labial
protraction by means of a multi purpose abdominal pump. Vid.
medd. Dan. naturalist. foren., 141: 81-96
PARRY, D.A. (1983): Labial extension in the dragonfly larva Anax imperator.
J. Exp. Biol., 107: 495-499
PETALURA vol. 1, 1993, p. 12
POPHAM, E.I. & BEVANS,
E. (1979): Functional morphology of the
feeding apparatus in larval and adult Aeshna juncea L.
(Anisoptera; Aeshnidae). Odonatologica, 8(4): 301-318
PRINGLE, J.W.S. (1963): Insect flight. Moscow. 178pp. (russian translation)
RÖDER, G. (1984): Morphologische Untersuchungen an Prätarsen von
Diptera und Mecoptera (Insecta). Diss., University of Nürnberg:
p. 393
RÖDER, G. (1986): Zur Morphologie des Prätarsus der Diptera und Mecoptera.
Zool Jahrb., Abt. Anat. und Ontog. Tiere, 144(4):
465-502
STEINMANN, H. (1967): A study of the cephalic measurement values of the species
of european Odonata families. Dtsch. entomol. Z., 14(3-4):
265-275
STORK, N.E. (1980): Experimental analysis of adhesion of Chrysolina polita
(Chrysomelidae, Coleoptera) on a variety of surfaces. J. Exp.
Biol., 88: 91-107
STORK, N.E. (1981a): The structure and function of the adhesive organs on
the antennae of male Harpocera thoracica (Fallen) (Miridae:
Hemiptera). J. Nat. Hist., 15(4): 639-644
STORK, N.E. (1981b): The adherence of beetle tarsal setae to glass. J.
Nat. Hist., 15(4): 583-597
STORK, N.E. (1983): A comparison of the adhesive setae on the feet of lizards
and arthropods. J. Nat. Hist., 17(6): 829-835
STRAUSFELD, N.J. & SEYAN,
H.S. & MILDE, J.J. (1987): The neck motor system of the fly Calliphora erythrocephala.
1. Muscles and motor neurons. J. Comp. Physiol. (A), 160:
205-224
TANAKA, Y. & HISADA,
M. (1980): The hydraulic mechanism of
the predatory strike in dragonfly larvae. J. Exp. Biol.,
88: 1-19
TILLYARD, R.J. (1917): The biology of Dragonflies (Odonata or Paraneuroptera).
Cambridge University Press: Cambridge. 396 pp.
WIGGELSWORTH, V.B. (1987): How does a fly ching to the under surface of a glass sheet?
J. Exp. Biol., 129: 363-367
ZAIKA, V.V. & VORONOVA,
I.A. (1977): Behaviour of dragonflies
in a lake-grassland of Western Sibiria. p. 82-105 in: Ethological
problems of sibirian insects ecology. University Novosibirsk
Press: Novosibirsk. (in russian)
Submitted 31. May 1993
Reviewed and accepted 20. July 1993
PETALURA vol. 1, 1993, p. 13
FIGURE LEGENDS
Fig. 1.)
Sketch of the head fixation system of odonates (dorsal view):
- 1 = Head free and mobile.
- 2 = Head in fixed position.
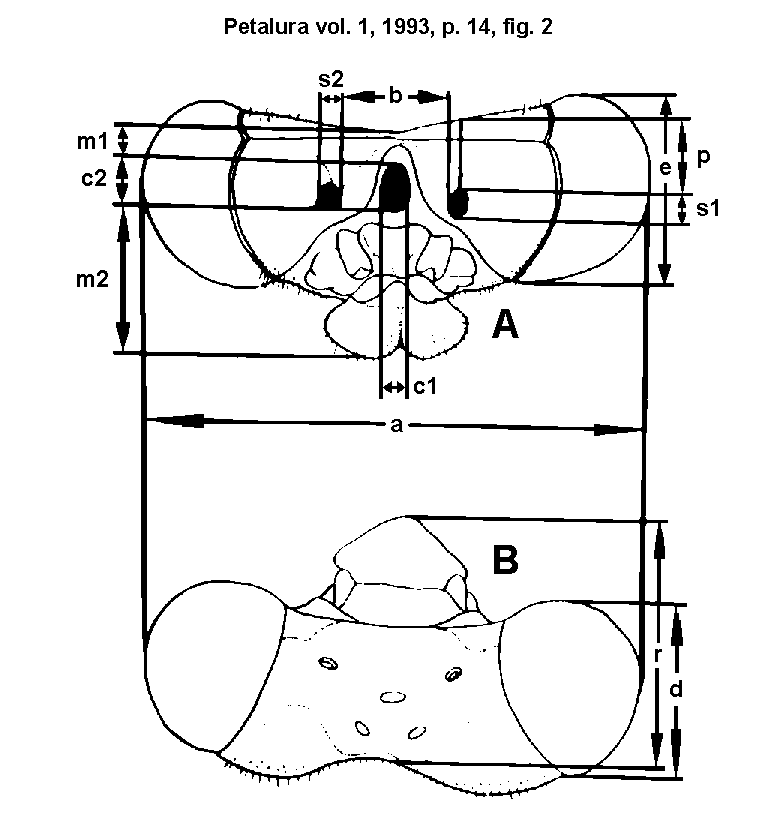
Fig. 2.)
Pattern of measurements used for cluster analysis:
a = total head width; b = distance between the medial borders
of the MFH (arrester base); c1 = maximum width of occiput (base
of the head-cephaliger joint); c2 = occiput height, d = eye length;
e = eye height; m1 = distance between the upper borders of the
head and the occiput; m2 = distance between the lower border of
the occiput and the mouthparts; p = distance between the upper
borders of the head and the MFH; r = distance between the posterior
and the anterior border of the head (head length); s1 = height
of the MFH; s2 = width of the MFH.
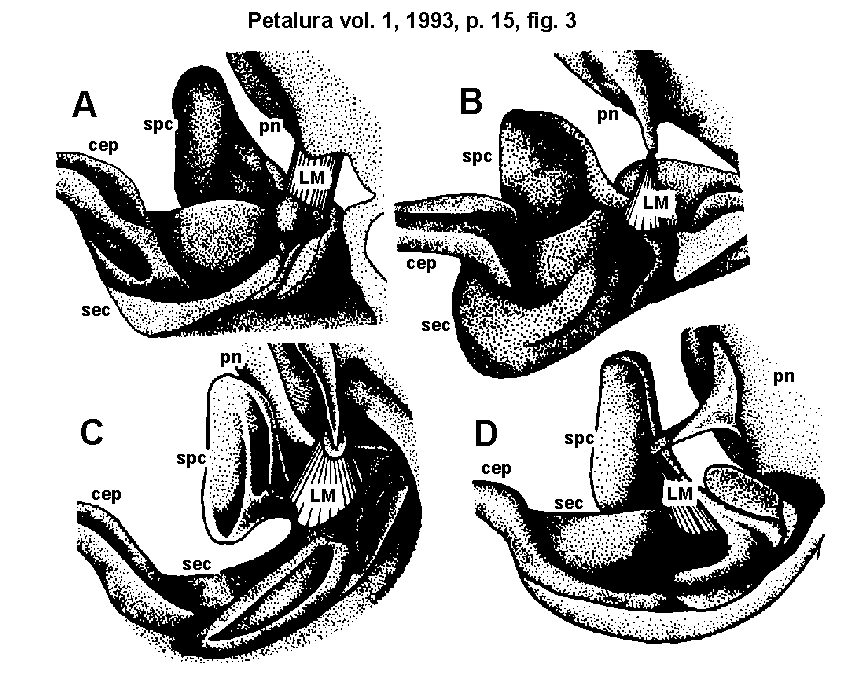
Fig. 3.)
Neck region of imaginal odonates (lateral view of the interior side):
- A = Cordulegaster annulatus Don.
- B = Cordulia aenea L.
- C = Aeshna mixta Latr.
- D = Gomphus flavipes Charp.
The lateral walls of the neck are formed by a pair of eucervical
sclerites (SEC). These SEC are transformed anteriorly into two
acute cephaligers (CEP). The SEC connect with the postcervical
sclerite (SPC) near the dorso-proximal edge of the SEC. The origin
of the SPC-abductor muscle (LM) is the pronotal apodem, its insertion
is the cuticular region between the SEC and the SPC.
Fig. 4.)
The cephaligers (CEP) are symmetrical relatively
to the longitudinal body axis. These processes converge anteriorly
to one point, which is the centre of gravity of the head. In this
point the head tentorium (TR) connects with the neck through a
pair of mobile joints. Drawing from Sympetrum sanguineum Mill..
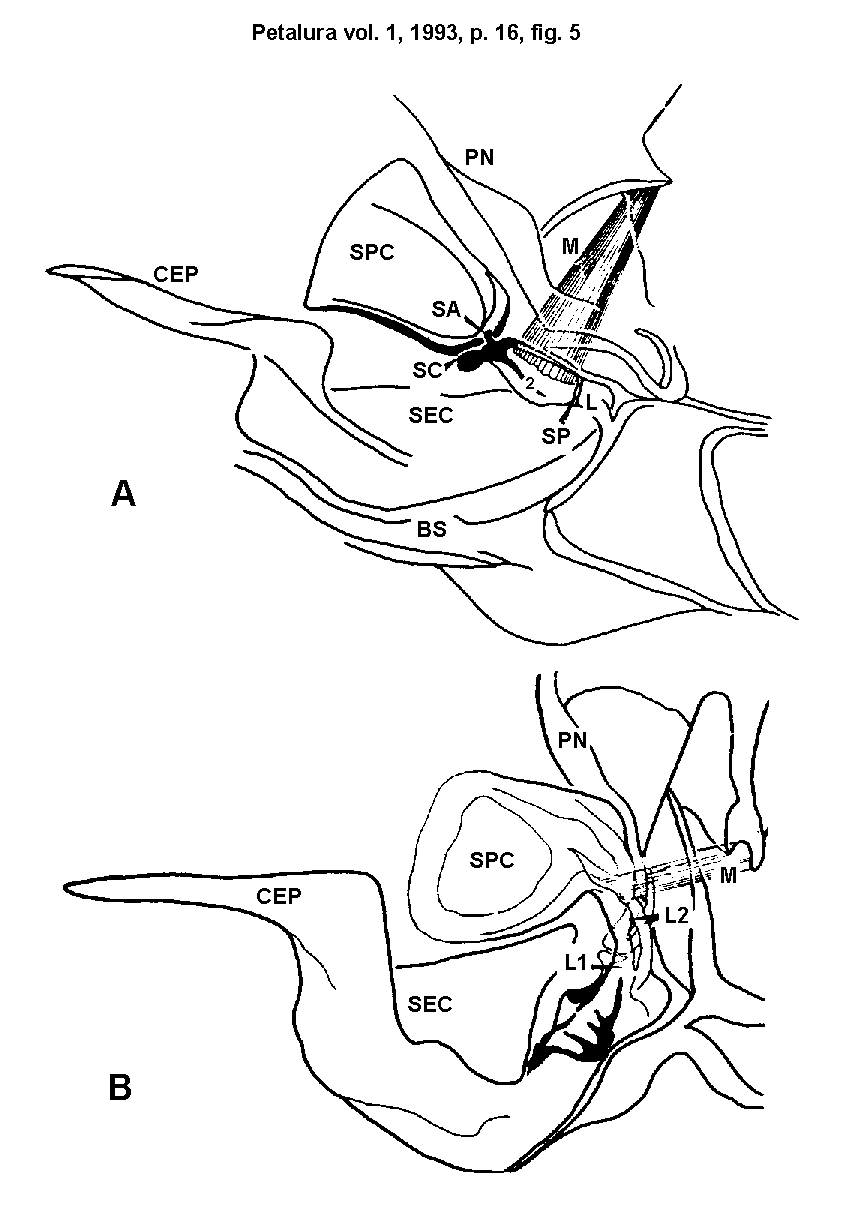
Fig. 5.)
Cuticular structures that achieve SPC-adduction:
- A = Sympetrum.
- B = Lestes.
Posteriorly the SPC and SEC are connected by parts of elastic
cuticula, which achieve the adduction of the SPC by a spring-mechanism.
The spring-effect is increased by a pair of hard cuticular "legs"
(L1, L2) of the SPC-SEC region. Besides, the SPC-"legs"
of Sympetrum do have two supplementary parts of elastic
cuticula (SA + SC, SP), - posteriorly and anteriorly.
Fig. 6.)
Cluster diagramm of the head characters:
The characters of the head capsule are well correlated with the
arrester base, but only weakly correlated with the size of the
microtrichia fields (s1, s2).
Fig. 7.)
Correlation of the arrester base (B), the base
of the head-cephaliger joint (C1) and the occiput height (C2)
with the head width (A).
Fig. 8.)
Correlation of the arrester base with the head
width in ...
- A = ZYGOPTERA: CO = Coenagrionidae; LE = Lestidae; CA = Calopterygidae.
- B = ANISOPTERA: AE = Aeshnidae; GO = Gomphidae; CO = Corduliidae; LI = Libellulidae.
Fig. 9.)
Free or fixed position of the head fixation system,
caused by arrester action in different behavioural situations:
- 1 = Tandem sitting.
- 2 = Roosting.
- 3 = Saccadic sagittal head movements during hunting and perching flight.
- 4 = Feeding.
- 5 = Flying with captured prey.
- 6 = Normal flight.
- 7 = Tandem flight.
Fig. 10.)
Morphological types of the MFH in Anisoptera:
- A = Gomphid-type.
- B = Anactid-type.
- C = Aeshnid-type.
- D = Libelluloid-type.
An enlarged arrester-base and the formation of a triangular contact
(two MFH and the cephaligers) increase the stability of the arrester joint.
 back
back  next
next

 E-Mail to the Editor: bechly@mac.com
E-Mail to the Editor: bechly@mac.com
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}