The anterior (lower) and median (upper) processes of the odonate
ovipositor are bilateral narrow lanceolate appendages which serve
to allow deposition of eggs in an opening in the plant surface,
in the soil, or in rotten wood. The functional morphology of the
odonate endophytic ovipositor has been studied by WEELE (1906),
TILLYARD (1917), QUENTIN (1962), and PFAU (1985, 1991) and the
neuroanatomy of central projections of zygopteran ovipositor mechanoreceptors
was investigated by GORB (1994a, b). It is known, that the inner
surface of the ovipositor leaves of some insects is covered by
microtrichia (MICKOLEIT, 1973; AUSTIN & BROWNING, 1981; FIELD
& AUSTIN, 1994). The morphological diversity of the microsculpture
of the inner surface of the ovipositor leaves in different odonate
phyla has not been studied previously.
We used dry collection material of following odonate species:
Anisoptera: Aeshnidae (Aeshna affinis v. d. Linden, Ae. cyanea Müll., Ae. juncea L., Ae. mixta Latr., Anaciaeshna isosceles Müll., Anax imperator Leach., Hemianax ephippiger Burm., Gynacantha nervosa Rambur, Aeshnophlebia anisoptera Selys); Anisozygoptera: Epiophlebiidae (Epiophlebia superstes Selys); Zygoptera: Calopterygidae (Hetaerina cruentata Rambur, Vestalis gracilis Rambur, Mnais pruinosa Selys, Matrona basilaris Selys, Calopteryx splendens Harris); Diphlebiidae (Diphlebia lestoides Selys); Chlorocyphidae (Libellago lineata Burm.); Polythoridae (Euthore fasciata Hagen); Lestidae (Sympecma annulata Selys, Lestes sponsa Hans., Archilestes sp.); Megapodagrionidae (Heteragrion chrysops Hagen, Philogenia cassandra Hagen, Oxystigma williamsoni Geijskes, Hypolestes trinitatis Gundlach); Perilestidae (Perissolestes romulus Kennedy); Coenagrionidae (Ischnura pumilio Charp., Nehalennia speciosum Charp., Erythromma najas Hans., E. viridulum Charp.); Protoneuridae (Epipleoneura lamina Williamson), Platycnemididae (Copera annulata Selys, Platycnemis pennipes Pall.); Pseudostigmatidae (Mecistogaster ornatus Rambur).
For scanning electron microscopy, the ovipositor leaves were washed in alcohol (70°), air-dried, mounted on holders, sputtered with gold and examined under a JEOL JSM-35 scanning electron microscope.
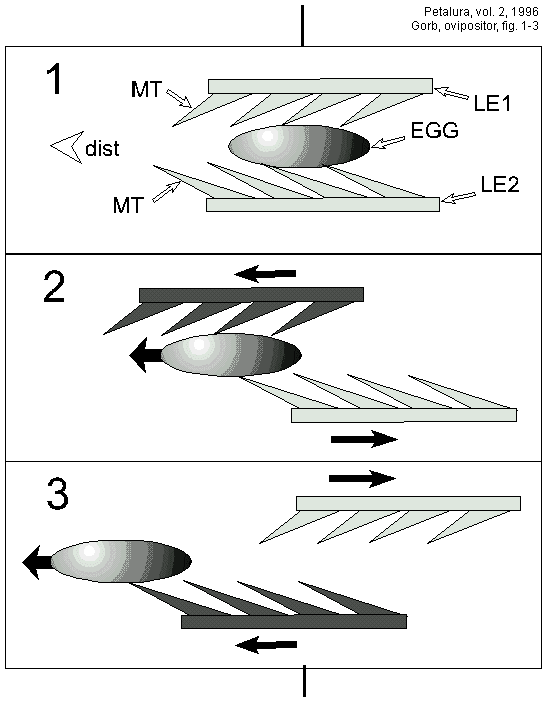
The microtrichia are situated on the inner surfaces of the leaves, and are directed postero-distally. Thus, the opposing movements of the leaves result in egg displacement in only one direction (distally). It may be suggested, that the observed structures facilitate the transport of eggs out of the ovipositor (Figs. 1-3).
Different odonate taxa have a different morphology and arrangement of the ovipositor microtrichia. The spectrum of the microtrichia types, which are present within a single species, may vary from one (A. imperator, Coenagrionidae) to four types (Ae. cyanea, P. romulus). Additionally, some species have a specific microsculpture in the proximal and marginal parts of each leaf and at the area of leaves' contact (sliding canals and sliding protuberances). Probably the microsculpture of contacting surfaces provides an increasing of ability to slide to one direction (anterior-posterior) and prevents sliding to another one (lateral-medial).
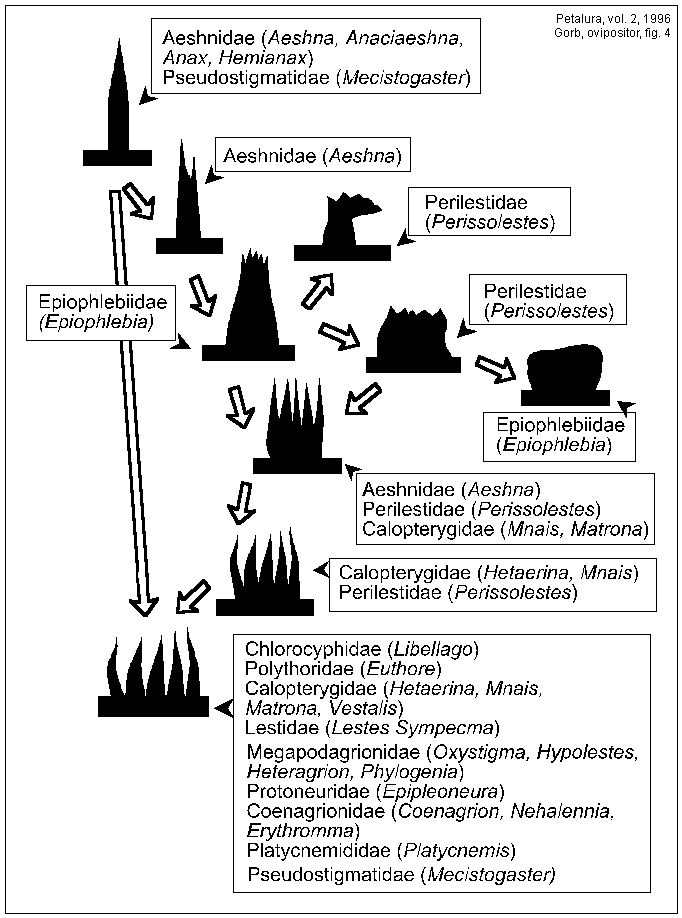
The spectrum of the modification of the structures on the inner surface of the ovipositor leaves is very wide. However, three main types of structures can be observed: (1) single sparsed microtrichia, which are present in Aeshnidae and on the upper leaf in Zygoptera; (2) comb-like crest compositions, which are present mainly on the LL (rarely on the UL) in Zygoptera; (3) plates of Anisozygoptera. The differences in microtrichia morphology and distribution between the upper leaf (UL) and lower one (LL) are most evident in Epiophlebia superstes.
It is difficult to draw a possible evolutionary path of the structures described, because all types of the microstructures have a lot of transitional forms. It is hypothesised that the evolution of the microstructures might have been directed from the single sparsed microtrichia through the microtrichia with shallowly divided crests to the comb-like microtrichia pattern. It is probable that the plate-like microsculpture of the UL of E. superstes have formed parallel to the comb-like microtrichia of Zygoptera. However the proposed path of possible evolution of the microtrichia (Fig.4) does not at all reflect species evolution, but only the evolution of microsculpture of ovipositor leaves. Actually, the opposite evolutionary direction might well be possible, since comb-like microtrichia are not only occurring in all three suborders of the Odonata, but also in other insect orders (MICKOLEIT, 1973; AUSTIN & BROWNING, 1981; FIELD & AUSTIN, 1994).
Mr. M. SUGIMURA (Tombo Ohkoku, Nakamura City, Japan), Dr. DeMARMELS
(Univ. Central Venezuela), Prof. Dr. M. SAMWAYS (Univ. Natal,
South Africa), Dr. S. DUNKLE (Gainesville, USA) Dr. S. BORISOV
(Inst. Zool., Dushanbe, Tadjikistan), and Dr. L. PRITYKINA (Paleontological
Inst., Moscow, Russia) kindly donated specimens. I wish to thank
Dr. M. LANDOLFA (Inst. Zool., Univ. Wien) for valuable discussion
on the general problem of the evolution of structure and function.
The poster presentation at the First Odonatological Symposium
of the Alps-Adriatic regional Community (Maribor, Slovenia,
3-7 July 1994) was supported by The International Science Foundation,
New-York, NY 10016, USA (travel grant #1521/1).
AUSTIN A.D. and T.O. BROWNING (1981) A mechanism for movements
of eggs along insect ovipositor. Int. J. Insect Morphol. Embryol.
10: 93-108.
GORB S.N. (1994a) Central projections of ovipositor sense organs in the damselfly Sympecma annulata Selys (Zygoptera, Lestidae). J. Morphol. 220:139-147.
GORB S.N. (1994b) Damselfly ovipositor leaves: microsculpture of the surfaces moving eggs and neuroanatomy of central projections of ovipositor sensilla. Abstr. Pap. 1st Odonatol. Symp. Alp.-Adriatic reg. Comm. Maribor, Slovenia, 3-7 July 1994: 20.
FIELD S.A. and A.D. AUSTIN (1994). Anatomy and mechanics of the telescopic ovipositor system of Scelio Latreille (Hymenoptera: Sceleonidae) and related genera. Int. J. Insect Morphol. Embryol. 23: 135-158.
MICKOLEIT G. (1973) Zur Anatomie und Funktion des Raphidiopteren-Ovipositors (Insecta, Neuropteroidea). Z. Morphol. Tiere. 76: 145-171.
PFAU H.K. (1985) Die eigentümliche Eiablage der Cordulegaster-Weibchen. Natur Museum. 115:77-86.
PFAU H.K. (1991) Contributions of functional morphology to the phylogenetic systematics of Odonata. Adv. Odonatol. 5:109-141.
QUENTIN St., D. (1962) Der Eilegeapparat der Odonaten. Z. Morph. Ökol. Tiere. 51:165-189.
TILLYARD R.J. (1917) The Biology of Dragonflies (Odonata or Paraneuroptera). Cambridge: University Press.
WEELE van der H.W. (1906) Morphologie und Entwicklung der Gonapophysen der Odonata. Tijdschr. Ent. 49:99-198.
Figs. 1-3: Principle schema showing how the microtrichia (MT)
of the inner surface of the ovipositor leaves produce distally-directed
egg movements. The microtrichia are always angled posteriorly.
The leaf moving distally (darkly shaded) carries an egg out, whereas
the leaf moving in the opposite direction (lightly shaded) does
not generate egg displacement. dist - distal direction, EGG -
an egg, LE1, LE2 - ovipositor leaves, MT - microtrichia.
Fig. 4: Hypothetical schema of the possible course of the evolution of microstructures on the inner surface of odonate ovipositor leaves.
back
next

 E-Mail to the Editor: bechly@mac.com
E-Mail to the Editor: bechly@mac.com
{kind=link}
{kind=link}